Знание демографических процессов развития вредителей — основа высокоэффективного управления их численностью

Тепличная белокрылка, паутинные клещи, трипсы, галловые нематоды и многие другие опасные вредители растений в теплицах отличаются огромной плодовитостью, быстротой индивидуального развития и смены поколений.
Из-за этих биологических особенностей даже при высоком уровне смертности (95-99%) в результате истребительных мероприятий порой оказывается возможным не сокращать их численность, а лишь сдерживать темпы ее прироста. Основа для радикального улучшения результатов защитных мероприятий видится в использовании знаний демографических закономерностей, по которым развиваются популяции фитофагов. Сложность их популяционной структуры (наличие возрастной и пространственной гетерогенности) обусловливает необходимость разработки глубоко продуманных интегрированных систем защиты растений, отвечающих, по меньшей мере, принципу «демографической адекватности».
ВВЕДЕНИЕ
Суть рассматриваемой в статье проблемы можно свести к понятной всем аналогии. Как можно расценить действия командира, который после боя, завершившегося большими потерями в живой силе противника, рапортует начальству об успехе, но при этом игнорирует данные разведки о стекании в район дислокации новых превосходящих вражеских сил? Ответ представляется очевидным. Однако в области защиты растений с подобными «рапортами» мы сталкиваемся, к сожалению, постоянно. Более ранние публикации автора на эту тему [3-7] большей частью неизвестны или недоступны практикам, поэтому для пользы дела будет не лишним высказаться вновь.
Начнем анализ с примеров простейших расчетов, в основу которых положены хорошо известные из литературы данные о биологии основных видов вредителей [1, 10 и мн. др.], бесконечная «война» с которыми заставляет тепличные хозяйства из года в год тратить немало средств. Уже сам факт этот свидетельствует о наличии постоянного притока свежих сил «противников». Итак…
«УРОК АРИФМЕТИКИ»
Если мы знаем, что средняя плодовитость одной самки тепличной белокрылки Trialeu rodes vaporariorum (далее – ТБ) около 130 яиц, соотношение полов ~ 1:1, а какое-то средство Х может вызвать гибель 90% имаго, это означает, что из каждых 100 особей 5 самок, которые избежали гибели, способны за 3 недели произвести в среднем 650 потомков. Если говорить об агентах биоконтроля ТБ, то среди них высоким летальным действием на имаго обладают, пожалуй, лишь некоторые штаммы энтомопаразитического гриба Lecanicillium muscarium*. Тщательно обрабатывая листья огурца суспензией, содержащей 55107 конидий/мл и поддерживая в первые 24-36 ч пос ле опрыскивания температуру 25±2 °С и, главное, близкую к 100% относительную влажность воздуха, через 7-10 суток можно уничтожить 90-93% особей имаго. Двукратное увеличение инфекционной нагрузки повысит смертность до 95-97%, а четырехкратное – до 97-99%. На первый взгляд, такие различия выглядят статистически недостоверными, а поэтому дополнительные затраты – неоправданными. Однако две избежавшие уничтожения самки могут обеспечить в следующем поколении прирост популяции в среднем в 2,6 раза, тогда как одна – в 1,3 раза. Следовательно, при почти одинаковой в последних двух случаях смертности различия в численности живых особей после обработки могут оказаться весьма ощутимыми. К сожалению, на практике достичь столь высокого уровня смертности очень трудно даже при использовании «убойных» химических препаратов, и это позволяет предвидеть, что дальнейшие события будут развиваться по более худшему сценарию. Впрочем, при отсутствии как индуцированной, так и естественной преждевременной гибели (обычно она не превышает 5-10%, но при очень высокой плотности может достигать 20-35%) из каждых 100 особей имаго 50 самок могут отложить в общей сложности более 6000 яиц, а порой и в несколько раз больше!
У обыкновенного паутинного клеща Tetranychus urticae (далее – ОПК) самки откладывают в среднем по 4 яйца в сутки, а всего (за 3 недели) – 80, но иногда и до 150. Использование против этого вредителя биохимического препарата фитоверм-М (2 г/л) в рекомендуемой концентрации рабочего раствора 0,1% при температуре 25±3 °С и высоком качестве опрыскивания через 4-7 дней обеспечивает гибель имаго на 88-92%. Из избежавших гибели 8-12 особей ~2/3 составляют самки. Таким образом, оставшиеся 6-8 самок вместе могут уже через 4 дня дать 100 потомков (т.е. почти мгновенно восстановить прежнюю численность), а за весь репродуктивный период – 480-640, но при особо благоприятных условиях – до 1200. При соотношении самок и самцов 3:1 (такое часто наблюдается в весенний период при умеренных температурах) возрастание численности может быть, соответственно, еще сильнее.
*Именно на основе этого гриба в свое время в СССР был разработан биопрепарат вертициллин против тепличной белокрылки (в настоящее время он отсутствует в «Списке пестицидов… 2009 г.» [24]), а в Великобритании – Mycotal®. До недавнего времени в литературе широко фигурировало другое название – Verticillium lecanii, но теперь молекулярно-генетическими методами доказана принадлежность гриба к самостоятельному роду, причем сам вид был раздроблен на три новых, одним из которых и является L. muscarium.
Самки западного цветочного трипса Frankliniella occidentalis (далее – ЗЦТ) живут около месяца, и могут отложить до 300 яиц. Следовательно, при гибели 95% особей 4-5 выживших самок (у этого вида доля самцов в популяции часто бывает ничтожной) могут произвести 1500 потомков. Даже при отсутствии резистентности использование фосфорорганических препаратов (на пример, актеллика) или баковых смесей препаратов на основе авермектинов (фитоверм-М) и неоникотиноидов (актара) позволяет уничтожить почти полностью личинок младших возрастов, но гибель пронимф, нимф (они большей частью уходят с растений в грунт) и имаго в лучшем случае достигает 85-95% со всеми вытекающими последствиями.
У галловых нематод (ГН) (близкие виды Meloidogyne inco g nita, M. javanica, M. arenaria) даже при уничтожении 98 самок из 100 оставшиеся в живых две особи в состоянии обеспечить прирост популяции в следующем поколении в 8 раз при обычной плодовитости 400 яиц. Но она может достигать порой и 2-3 тыс. яиц, и тогда популяция, несмотря на столь значительную элиминацию, может возрасти в следующем поколении потенциально в 60 раз; а при снижении смертности до 90% – в 300 раз! Разница, как мы видим, огромная. Необходимо отметить, что примерно так и происходит в тепличных хозяйствах после пропаривания грунтов, при качественном выполнении которого можно уничтожить ГН практически полностью в толще до 25-30 см, однако благодаря не большой доле инвазионных личинок, сохраняющихся глубже, численность вскоре начинает неизбежно нарастать. При несоблюдении регламентов (экономия пара, сокращение экспозиции) взрыв численности происходит раньше и в больших масштабах. Последующее внесение в грунт перед посадкой рассады нематицидного препарата фитоверм-П (8 г/кг) в дозе 94 г/м2 [24] позволяет гарантированно предотвратить потери урожая только за счет того, что растения до плановой уборки урожая просто успевают ускользнуть от критического нарастания численности ГН, которое происходит несколько позже [2, 6].
Из представленных примеров следует, что контроль численности указанных выше вредителей (и многих других видов с аналогичной жизненной стратегией, обладающих огромным потенциалом воспроизводства в теплицах) объективно очень труден. Тем не менее, это не должно служить оправданием для того, чтобы выдавать желаемое за действительность, особенно, когда мы сталкивается с молниеносными темпами роста численности вредителей. Попытаюсь обосновать эти критические слова.
Согласно традиционной трактовке [11], биологическая эффективность применения пестицида – это результат, который может выражаться показателями гибели, снижения численности вредных организмов, степени повреждения ими защищаемых растений. Однако в подобных определениях явно не учитываются ситуации (не столь уж редкие), когда одновременно с гибелью происходит опережающий рост численности за счет интенсивного размножения и, отчасти, иммиграции (но на первых порах именно от нее во многом зависит дальнейшее фитосанитарное благополучие). Следовательно, в таких случаях мы можем лишь сдерживать темпы прироста численности, но не снижать ее. Как видно из приведенных примеров, небольшая (всего в несколько процентов) разница в гибели может кардинально изменить ситуацию. Уже этого достаточно, чтобы признать ис ключительно неадекватной широко известную и применяемую формулу оценки биологической эф фективности (БЭ):
БЭ = Nm / Ni х 100, (1)
где Ni – исходная относительная численность, Nm – численность погибших особей в результате проведения истребительного мероприятия.
Надо понимать, что это – всего лишь показатель смертности, который, бесспорно, очень важен на этапе скрининга инсекто-, акаро-, нематоцидных веществ, разных видов био-агентов или их внутривидовых форм (рас, штаммов и т. д.) для выявления их потенциала, но при оценке положения дел в производственных условиях он должен иметь лишь подчиненное значение. Приведенный в формуле (1) способ расчета может использоваться для оценки БЭ лишь в тех случаях, когда отсутствует одновременный прирост численности. Но это условие выполняется даже далеко не во всех лабораторных опытах. Например, можно строго изолировать в садке 100 тлей, обработать их суспензией конидий гриба Lecanicillium longisporum* или подсадить несколько особей хищников-афидофагов (скажем, клопа Macrolophus nubilis) и через несколько суток обнаружить, что, несмотря на наличие смертности, сумма живых и погибших превышает 100. И если в полевых условиях, когда мы имеем дело с вредителями, развивающимися в одном поколении, по формуле (1) еще можно судить с большой натяжкой об успешности, то в отношении подавляющего большинства тепличных вредителей расчеты по ней лишь вводят практиков в заблуждение. В рекламных инструкциях по применению самых разных препаратов часто можно увидеть слова о высокой биологической эффективности, достигающей 90-98% при использовании в каких-то рекомендуемых нормах. Прочитав такую информацию, агроном добросовестно применяет подобное средство, а потом наблюдает, что через 5-10-15 дней вредителя вовсе не становится намного меньше, как должно быть! Видя такую явную нестыковку, он имеет полное право предъявить иск за недобросовестную рекламу, и правда окажется на его стороне, ибо его прямая должностная обязанность – заботиться о получении максимально возможного урожая, который зависит от численности живых вредителей, а не погибших.
*На основе этого вида, который ранее тоже указывался в литературе под названием Verticillium lecanii, в Великобритании для борьбы с тлями был разработан коммерческий биопрепарат Vertalec®.
В приведенных выше четырех примерах были описаны весьма упрощенные ситуации. Но в действительности лишь в первое время, когда в теплицах появляются особи-основатели вредителей, их популяции представлены, как правило, одной стадией раз вития. Но вскоре происходят разительные изменения – популяции приобретают определенную и возрастную и пространственную структуру. На этих вопросах следует остановиться подробнее, поскольку именно их недооценка ведет ко многим просчетам.
В действительности всё сложнее
Важным моментом является то, что все беспозвоночные, будучи пой-килотермными (холоднокровными) организмами, очень чутко реагируют на изменения гигро-термических условий. А они существенно отличаются в разных местах теплиц и, более того, в ярусах листьев, расположенных на разной высоте. Поэтому у особей на листьях нижнего и среднего ярусов многие биологические характеристики (плодовитость, темпы развития и т. д.) оказываются одними, а на листьях верхнего (здесь температура выше, влажность ниже, освещенность лучше) - другими. Особи на растениях, находящихся по периметру теплиц и вдоль центральной дорожки, тоже имеют иные демо графические особенности по сравнению с теми, которые живут в глубине массива растений. То же самое касается ГН, поскольку с глубиной меняются многие характеристики тепличных грунтов: температура, влажность, плотность, степень аэрации и т.д. Постепенно из-за различий в темпах развития разных особей практически у всех вредителей в теплицах происходит наложение во времени одних стадий развития на другие, а позже – и поколений. Но основную лепту в эти процессы вносит все же другая общая особенность биологии – большая или меньшая продолжительность периода яйцекладки.
У ТБ первичные очаги чаще всего формируются в связи с залетом (заносом) имаго. Таким образом, очень короткий период популяция (субпопуляция) представлена именно этой стадией. Но уже спустя 4-5 дней, когда каждой самкой может быть отложено по 20-75 яиц, имаго становится лишь 2,5-10% от общей численности. После этого дней через 10, когда будет отложено еще по 40-150 яиц, доля имаго сократится до 1-3%, но появится новая балансовая составляющая – порядка 15-20% будет приходиться на личинок, отродившихся из первых яиц. Спустя еще неделю самки-основательницы начнут естественно отмирать по старости, но на замену им к этому времени или чуть позже начнется выход новых самок из первых потомков, которые вскоре приступят к яйцекладке, и т.д.
У ГН способностью к долгому выживанию в отсутствии подходящих растений обладают только инвазионные личинки второго возраста. Именно они играют роль основательниц. При этом одни особи имеют шансы внедриться в корни довольно быстро после посадки рассады на постоянное место, другие могут проникнуть в них через полтора-два месяца и позже, т. е. когда их «старшие сестры» могут стать даже «бабушками». Усложнение популяционной структуры в значительной мере связано с тем, что период яйцекладки растянут на 2-3 месяца (в среднем – около 75 дней).
Важно отметить, что все эти процессы только кажутся очень сложными, хаотичными и непрогнозируемыми, но это не так. И хотя соотношение разных стадий развития находится в постоянном движении «вверх – вниз», прослеживается устойчивая тенденция к относительной стабилизации.
Поскольку у всех этих вредителей самки за период жизни производят сотни потомков, со временем в популяциях яйца начинают значительно преобладать над другими стадиями.
Когда из этих яиц начнется массовый выход личинок, лидирующие позиции, однако, они не займут, т. к. из «старых» нимф выведутся имаго, которые будут в опережающих темпах откладывать сотни новых яиц. В связи с этим в «зрелых» популяциях доля имаго практически всегда минимальна, а доля личинок разных возрастов (включая нимф) занимает промежуточные значения. В общем виде эту закономерность можно представить так:
Nяиц > Nличинок > Nимаго.
У ТБ «среднестатистическая» популяция примерно такая: яйца – 55-95, личинки 1-3 возрастов и нимфы – 20-40, имаго – 0,5-5% от общей численности. У ОПК весной в период нарастания численности возрастная структура выглядит следующим образом: яйца – 60-80, личинки и нимфы – 10-35, имаго – 6-10%. У ГН: яйца – 40-96, личинки – 4-55, имаго – 0,2-5% [6]. Знание подобных закономерностей может дать очень многое.
Так, если мы знаем, что у ТБ личинки составляют всего 1/4-1/5 часть популяции, это означает, что даже при 100%-ной их гибели никакой сугубо ларвицидный агент будет не в состоянии сократить разновозрастную популяцию. Именно по этой причине около 30 лет назад потерпел фиаско масштабный эксперимент во многих тепличных хозяйствах страны, когда пытались бороться с этим вредителем с помощью узкоспециализированных энтомопаразитических грибов рода Ascher sonia, поражающих только личинок 1-3 возрастов. На этом примере еще раз можно убедиться в порочности использования в качестве показателя «высокой» БЭ действительно высокой гибели лишь одной стадии развития насекомых-вредителей.
В связи со сказанным будет уместно показать наглядно судьбу популяции ТБ после совместного применения в баковой смеси спор двух видов энтомопаразитических грибов – Lecani cil lium muscarium и Beauveria bassiana*. В отличие от грибов рода Aschersonia они в той или иной мере могут уничтожать все стадии развития [1, 3]. Представленная на рис.1 схема-матрица дает ясное представление, какие сложные и быстротечные процессы могут произойти за 8 дней после обработки этими биоагентами растений огурца в теплице при температуре 25±2 °С.
*На основе этого гриба, поражающего многих насекомых из разных отрядов (в частности трипсов, личинок минёров и отчасти тлей и др.), в нашей стране уже давно был разработан биопрепарат боверин. Однако в последние годы он исключен из Списка рекомендованных препаратов, поскольку не прошёл процедуру перерегистрации.
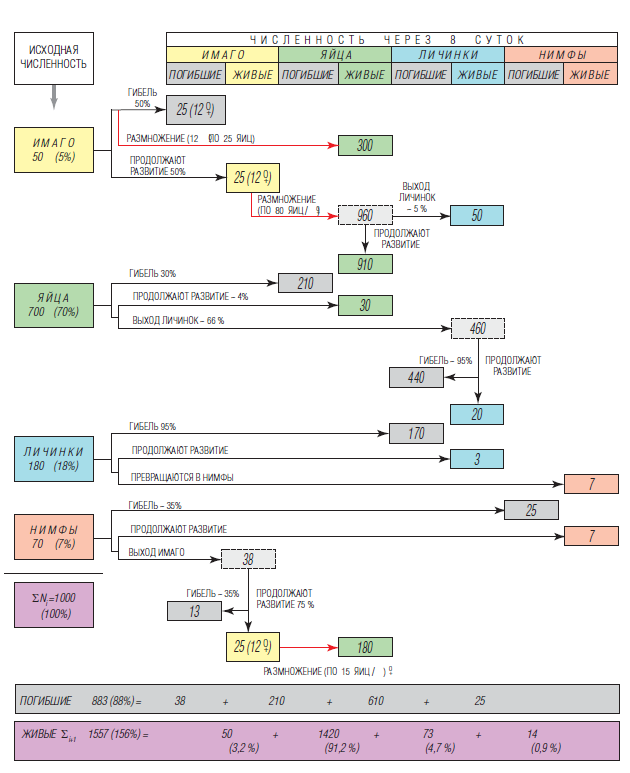
Здесь необходимы некоторые пояснения к аналитической модели. Предположим, что исходная плотность популяции 1000 особей, а возрастная структура достаточно типичная: имаго – 5, яйца – 70, личинки – 18, нимфы – 7%. В модель заложена вполне реальная плодовитость самок – 200 яиц (т.е. по 8-10 яиц в сутки), но одновременно – почти предельные показатели гибели разных стадий, на которые можно рассчитывать при обработке листьев суспензией, содержащей по 75х106 конидий/ мл каждого из возбудителей, а также эмульгатор Сильвет®Голд (0,03%), технический глицерин (0,3%) и мочевину (0,4%) в качестве активаторов прорастания конидий. Обращаю внимание, что в модели указана гибель имаго на уровне 50%, хотя выше говорилось (см. п. 2.1), что при таких дозах она может превышать 90%. В этом нет ошибки. Дело в том, что не менее 50% взрослых особей сосредоточено на 3-7 листьях верхнего яруса, где практически невозможно создать высокую влажность, необходимую для прорастания конидий и успешного заражения. В то же время на листьях среднего и нижнего ярусов гибель имаго действительно может достигать 93-97%. Важно отметить, что зараженные, обреченные на гибель самки, тем не менее, еще 2-4 суток тоже продолжают вносить свою лепту в прирост популяции. Соответственно самки, избежавшие контакта с инфекцией, имеют возможность размножаться все дни в полную силу (по 10 яиц/сутки). Хотя овицидная активность этих энтомопаразитов невелика, это компенсируется высокой смертностью отрождающихся личинок-бродяжек, которые захватывают на себя много инфекционных конидий, передвигаясь по поверхности листьев в поиске места для присасывания. Нимфы гораздо менее уязвимы для инфекции, но около 1/3 их все же может погибнуть в основном благодаря действию гриба B. bassiana. Выхо дящие из них имаго тоже отчасти подхватывают инфекцию и вскоре гибнут, а те из них, которые быстро мигрируют в верхний ярус растений, избегают этой участи и даже успевают через 8 суток дать небольшое потомство. Обращаю внимание на выделенные красным цветом три стрелки: это и есть те самые «магистрали», где нас подстерегает опасность.
Что мы имеем в итоге? Совокупную гибель на всех стадиях развития 88%. Но живых особей остается не 12%, а почти в 1,6 раз больше, чем было! Конечно, здесь можно утешаться тем, что в отсутствии обработки ситуация была бы хуже – при данных параметрах плодовитости, возрастной структуры и нулевой смертности популяция в контроле возросла бы через 8 суток в 3,5 раза. Но гораздо правильнее сопоставить полученный результат с исходной численностью по одной из двух формул (они равнозначны):
БЭ = (1 – ƩNi+1/ SNi) x 100, (2)
или
БЭ = (SNi – SNi+1)/SNi x 100. (3)
Здесь: ƩNi – исходная суммарная относительная численность вредителя на всех стадиях развития, ƩNi+1 – совокупная численность особей после истребительного мероприятия [3, 14].
В нашем примере БЭ1 = (1 – 1557/1000 ) x 100 = – 55,7 %. Это совершенно правдивый показатель! Минус надо воспринимать как общую оценку проведенной работе, в результате которой именно на столько процентов не уменьшилась, а возросла численность, и, значит, ситуация остается опасной и необходимо продолжать бороться с вредителем, используя либо прежний арсенал, либо что-то иное.
В этой связи интересно построить следующую модель для прогноза возможных результатов от повторного применения тех же биоагентов, заложив в нее все прежние условия, а в качестве исходной структуры популяции взяв числовые характеристики из нижней графы первой модели (ЖИВЫЕ ƩNi+1): имаго – 50 экз. (3,2%), яйца – 1420 (91,2%), личинки – 73 (4,7%), нимфы – 14 (3,2%). Из-за ограниченности объема статьи схема не приводится, но читатели могут самостоятельно сделать расчеты и убедиться, что в этом случае произойдет уже небольшой (всего на 8 %) спад численности по сравнению с той, которая была после первой обработки. Это объясняется прежде всего тем, что в структуре популяции доля имаго здесь меньше, чем была первоначально: 3,2 вместо 5%. Однако говорить об успехе вновь не приходится: БЭ2 = (1 – 1429/1000 ) x 100 = – 42,9%, т.е. численность все равно выше исходной более чем на 40%. Точно так же можно оценить и возможные результаты последующих обработок и увидеть, что третья, наконец, позволит получить небольшую эффективность со знаком «+»: БЭ3 = (1 – 867/1000) 5 100 = +13%. А четвертая приве дет к сокращению численности примерно в 2 раза – до ~500 особей ТБ.
В 80-х годах минувшего столетия, когда во многих тепличных комбинатах страны пытались бороться с ТБ с помощью этих видов биоагентов (чаще использовали лишь один вид – L. mus carium), более-менее приемлемые результаты снижения численности в 8-12 раз получались только после 6-15 последовательных обработок, что подтверждает адекватность моделей. Очевидно, что с экономических позиций это далеко не лучший путь.
Аналогичные аналитические модели можно построить и для оценки тенденций изменения численности других вредителей при использовании против них разных средств. Смею заверить, что в большинстве случаев они не будут кардинально отличаться, отражая тем самым реальное, из года в год повторяющееся положение дел в тепличных хозяйствах.
Один пример, однако, будет не лишним рассмотреть вкратце. Выше (см. п. 2.2) уже говорилось, что при использовании фитоверма-М в 0,1%-ной концентрации у ОПК гибель имаго составляет около 90%, а прирост численности может быть в 5-10 раз и более. В то же время при использовании препарата в 5-кратной «трипсовой» дозе смертность имаго повышается на 5-6%, и благодаря этому выживают не 6-8, а 1-3 самки. Поэтому следующее поколение возрастает не столь сильно. Поскольку популяции ОПК имеют сложную возрастную структуру, добиться радикального снижения численности даже при такой дозе можно не менее чем 3-6 обработками. Это во многом будет зависеть от соотношения разных стадий развития к моменту проведения защитного мероприятия, а точнее, от большей или меньшей доли самок, а также от плодовитости в конкретный период. Если же придерживаться регламентированной концентрации рабочего раствора 0,1% и проводить обработки к тому же с интервалом 20 дней (можно представить, какой прирост будет за 3 недели!), как указано в официальном документе, то борьба с этим вредителем приобретает гораздо более затяжной характер. Это яркий пример, к чему может вести недооценка демографических особенностей вредителей и использование неадекватных формул при регистрационных испытаниях.
ЧТО СЛЕДУЕТ ИЗ СКАЗАННОГО?
Напрашивается вывод: чем больше на данном временном срезе в структуре популяции любого вредителя доля имаго (обычно в самый начальный период) и чем выше продуктивность самок (обычно тоже в более ранние сроки при меньшей плотности), тем стремительнее будет расти численность и тем более сильные и быстродействующие средства нужно применять.
Надо ясно понимать и то, что даже объективно оцененная БЭ имеет лишь относительный характер. Существует огромная разница между выживанием на одном растении в среднем одной особи из 20 или 5 тыс. из 0,1 млн. (у ТБ такая плотность в самом деле бывает в летний период при отсутствии действенных мер борьбы; в таких случаях на 1 см2 листовой поверхности можно учесть в среднем 7-20 экз.), хотя в обоих случаях БЭ = 95%. Поэтому повторю, что гораздо важнее знать не количество выбывших из популяции особей (сколько бы их ни погибло – пусть даже 99,9%), а живых, наносящих вред растениям.
Быстрота, с которой происходит рост численности многих вредителей в теплицах, – важнейший момент, который начисто обесценивает и значение при выборе сроков проведения защитных мероприятий широко используемого критерия – экономического порога вредоносности, хотя знать это непременно надо. Так, для ТБ указываются пороги: на культуре огурца – в среднем 40 экз. разных стадий на 1 лист, а на томате – 10 экз. Однако если сегодня мы зафиксируем, например, 20 особей, из которых будет только одна самка, то через 2 суток можно ожидать уже удвоения популяции. Поэтому меры должны приниматься незамедлительно сразу после обнаружения первичных очагов.
Из представленных данных следует: чтобы произошло хотя бы малейшее снижение численности вредителей, необходимо, чтобы одна выжившая самка из 100 особей приносила менее 100 потомков, 5 самок – в среднем менее 20 потомков каждая, 10 самок – менее 10 и т.д. Но для того чтобы можно было говорить о действительно высокой БЭ, «желательная» продуктивность фитофагов должна быть еще значительно ниже. Вот в этом и состоит главная сложность! Получается, что, несмотря на наличие каких-то резервов в увеличении доз, кратности обработок, в качестве опрыскивания (или внесения препаратов в тепличный грунт), все эти меры не могут служить адекватным ответом. К тому же нельзя экстраполировать данные лабораторных экспериментов на реальные производственные условия, ведь здесь, увы, рассчитывать на достижение 100%-ной гибели не приходится, но во многих слу чаях по объективным причинам и 90% – трудно досягаемая планка.
Таким образом, остается, по-видимому, единственный резерв – как-то дополнительно воздействовать на собственно плодовитость. Об этом уже давно писали Вронских и Булыгинская [9], но, к сожалению, эта мысль пока не получила должного практического воплощения.
Здесь уместно вспомнить японский препарат апплауд, который ныне не фигурирует в числе разрешенных средств, однако долгое время он был одним из наиболее эффективных против ТБ. Это объясняется тем, что его действующее вещество (бупрофезин), являющееся регулятором синтеза хитина, оказывает не только инсектицидное действие на личинок в период линьки (нарушение метаморфоза), но и приводит к откладке самками нежизнеспособных яиц.
Тот факт, что плодовитость растительноядных беспозвоночных имеет большие различия не только на разных видах, но и сортах (гибридах) растений, известна вообще-то давно (немало примеров представлено в книге [1]). У ГН на устойчивых гибридах томата могут меняться многие биологические (демографические) характеристики: удлиняться период развития, увеличиваться доля самцов в популяции, уменьшаться плодовитость и др., что ведет и к изменениям в структуре популяций. Вместе с тем необходимо обратить внимание, что задача многочисленных селекционных работ на устойчивость к ГН – прежде всего, получить растения, отличающиеся способностью полноценно развиваться и давать высокий урожай, несмотря на наличие вредителя.
Однако не лишен перспективности и целенаправленный отбор генотипических линий тепличных культур по признаку резкого снижения плодовитости как ГН, так и других фитофагов, если, конечно, этот полезный признак не будет одновременно сказываться негативно на вкусовых качествах, урожайности, лежкости плодов и т.д.
Но есть и другой путь, который в общем-то хорошо известен, но не нашел пока широкого применения (по-видимому, именно из-за того, что основным рычагом воздействия на вредителей традиционно считается их массовое уничтожение). Речь идет об актуальности продвижения в практику защиты растений в теплицах веществ с репеллентной и антифидантной активностью, благодаря которым вредители вынуждены затрачивать много энергии на поиск более подходящих растений, и не могут в полной мере реализовать свой биотический потенциал.
При сплошной обработке теплиц такими веществами с сильным и пролонгированным действием фитофаги вообще могут лишиться в замкнутом пространстве возможности продолжить свое развитие. Здесь заслуживают упоминания данные исследований, проводимых во Всероссийском НИИ защиты растений (ВИЗР, С.-Пе тербург). Так, в работе сообщается, что однократной обработки семян томата регулятором роста иммуноцитофитом (он есть в «Списке… 2009 г.») было достаточно, чтобы сделать кормовые растения непривлекательными для питания и яйцекладки ТБ. Аналогичное действие на ТБ проявили вещества метилсалицилат и метилжасмонат. Последний является известным активатором вторичного метаболизма растений; он показал высокие результаты и в отношении ЗЦТ: обработка растений томатов способствовала снижению численности потомства более чем на 80%.
Хорошо известно, что весьма сильной репеллентностью в отношении ТБ, тлей и многих других вредителей обладают, например, алкалоиды, содержащиеся во всех частях растения настурции большой Tropaeolum majus [1]. Но во многих случаях экстракты из растений обладают и комбинированным действием. Так, при изучении действия на ЗЦТ этанольного экстракта из надземной части растения Silene sussamyrica, произрастающего в Киргизии, было показано как прямое инсектицидное действие на личинок при их линьке, так и репеллентное в отношении взрослых самок, благодаря чему численность потомства снижалась примерно на 60%.
В последнем опубликованном «Списке…2009 г.» приводится не нашедший пока широкого применения в закрытом грунте препарат сочва на основе водорастворимых веществ, образующихся при пиролизе древесины. Этот репеллент в 0,3%-ной концентрации при опрыскивании растений огурца в течение нескольких дней «будоражит» имаго ТБ, тем самым не позволяя самкам приступить к яйцекладке.
Все больше накапливается данных о том, что аналогичным действием обладают и некоторые микроорганизмы. Так, после обработки листьев огурца культурой гриба Trichoderma ligno rum (современное название – T. viride) тли прекращали на них питание и падали на почву. Аналогичное (хотя и не длительное) действие было экспериментально подтверждено позже и в отношении ТБ при использовании одного из штаммов гриба T. harzianum: об работка суспензией с титром 108 конидий/мл приводила к тому, что в течение двух-трех суток имаго избегали садиться на опрыснутые листья огурца и не могли отложить на них яйца [Борисов, неопубликов. данные].
Чтобы было понятнее, какое огромное значение может иметь использование тех или иных веществ, сокращающих плодовитость, вернемся к аналитической модели, представленной на рис. 1. Если сохранить в ней прежний уровень инсектицидной активности двух видов паразитических грибов, но дополнить их действие гипотетическим агентом Y, способным снизить на всех трех «красных магистралях» плодовитость самок ТБ на 80%, то здесь уже после одной обработки численность уменьшится, по крайней мере, в 2,5 раза, а не возрастет, как было в первом случае.
Эти данные ясно показывают и предсказывают, что многие агенты биоконтроля, использование которых в «чистом» виде далеко не всегда дает должные результаты, при грамотном продуманном сочетании с другими средствами вполне могут занимать достойное место в интегрированных системах. Вместе с тем, если представленные выше аргументы послужат дополнительным стимулом для широкого внедрения в практику тепличного растениеводства разработок по линии сдерживания как такового размножения вредителей (т.е. с помощью репеллентов, антифидантов, гормональных препаратов, индукторов устойчивости и т.д.), то очень не хотелось бы, чтобы их использование стало рекламироваться в качестве очередной панацеи.
ЗАКЛЮЧЕНИЕ
Подводя итоги, необходимо сказать следующее. Борьба с вредителями (разумеется, и с болезнями тоже) приобретает осмысленность и становится по настоящему эффективной, когда она перестает уже быть борьбой, а переходит в иное качество – управление (в смысле английского слова control, а не management; отсюда термин biocontrol) популяциями. Раз работкой общей теоретической основы всякого управления сложными динамическими системами и процессами занимается особая научная дисциплина – кибернетика, одно из определений которой звучит исключительно в тему: «искусство обеспечения эффективности действия». В числе основоположников кибернетики был выдающийся английский ученый У.Р. Эшби, сформулировавший фундаментальный закон, известный как принцип необходимого разнообразия (Ash by’s Law of Requisite Variety). Говоря простым языком, этот закон гласит, что в любой сфере практического управления чем-либо или кем-либо сложность можно победить, как минимум, тем же уровнем сложности, но иногда – и большим, т.е. имея еще и определенный «запас прочности». Эквивалентные положения были независимо сформулированы и в других областях, связанных с вопросами управления. Так, в теории игр важнейшим постулатом является следующий: чтобы выиграть, необходимо иметь больше доступных ходов, чем ваш противник.
Применительно к разбираемой в данной работе проблеме это означает: чтобы уверенно выигрывать у вредителей, нужно умело учитывать в практических действиях сложность их популяционной структуры. Для краткости этот принцип был назван «демографической адекватностью» [6], но, по сути, это лишь следствие из законов более высокого ранга. Нет ничего удивительного в том, что, действуя вразрез им, сегодня мы имеем пока то, что имеем: бесконечную дорогостоящую «войну».
Автор: Б.А. Борисов, заведующий лабораторией микробиологических средств защиты растений ООО «АБТ»
СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ
1. Ахатов А.К., Ижевский С.С., Мешков Ю.И., Борисов Б.А., Волков О.Г., Чижов В.Н. Вредители тепличных и оранжерейных растений (морфология, образ жизни, вредоносность, борьба) / Под ред. А.К.Ахатова и С.С.Ижевского. М.: Товарищество научных изданий КМК, 2004. 307 с.
2. Березина Н.В. Сравнение способов обеззараживания грунта против комплекса галловых нематод в защищенном грунте (АОЗТ «Матвеевское», 2001-2002 гг.) // Интегрированная защита растений и элементы технологии в тепличных комбинатах Российской Федерации: Сб. докл. III и IV Всеросс. семинаров повышения квалификации специалистов по защите растений и агрономов-технологов. М.: Агроконсалт, 2004. С. 155-160.
3. Борисов Б.А. О необходимости учета структуры популяций вредителей и фазовой специфичности энтомопатогенов при выборе микробиологических средств защиты растений // Производство и применение грибных энтомопатогенных препаратов. М.: ОРИСО Главмикробиопром, 1985. С. 27-31.
4. Борисов Б.А. Оценка биологической эффективности средств защиты растений (на примере управления численностью тепличной белокрылки) //Микробиологические средства защиты растений: Сб. науч. тр. / ВАСХНИЛ, Сиб. отд. Новосибирск, 1986. С. 91-94.
5. Борисов Б.А. Экологически безопасная защита тепличных растений от галловых нематод: краткий очерк проблемы // Аграрная Россия. 1999. № 3. С. 35-42.
6. Борисов Б.А. Демографическая адекватность — важнейший принцип выбора наиболее эффективных способов контроля численности галловых нематод рода Meloidogyne // Тр. Центра паразитологии Института проблем экологии и эволюции им. А.Н. Северцова РАН. Т. 45: Сис тематика и биология паразитов / Под ред. С.А. Беэра. М.: Наука, 2008. С. 238-281.
7. Борисов Б.А., Ахатов А.К. Борьба с оранжерейной белокрылкой: анализ трудностей и поиск рациональных путей // Защита растений. 1991. № 9. С. 6-10.
8. Борисов Б.А., Березина Н.В., Мешков Ю.И. Биохимические препараты серии «Фитоверм»: применение против паутинных клещей рода Tetranychus в защищенном грунте // Гавриш. 2000. № 2. С. 18-25.
9. Вронских М.Д., Булыгинская М.А. Зависимость эффективности новых методов борьбы от биологического потенциала вредных видов насекомых // Бюлл. ВИЗР. 1981. № 51. С. 39-42.